Петр Олексенко. Южноазиатское происхождение человечества. Краткое эссе книги С. Оппенгеймера «Изгнание из Эдема»

В своей книге «Изгнание из Эдема. Хроники демографического взрыва» (OUT OF EDEN. THE PEOPLING OF THE WORLD, 2003) английский исследователь, медик, генетик, палеоантрополог Стивен Оппенгеймер (Stephen Oppenheimer) на основе генетических данных аргументированно излагает гипотезу о том, что жители Евразии имеют южноазиатское происхождение.

Стивен Оппенгеймер (Stephen Oppenheimer, род 1947), английский исследователь и автор книг по доисторическому периоду человечества. В его книгах вопросы генетики рассматриваются в связке с археологией, лингвистикой и фольклором.
В основных географических пунктах Y-хромосомы служат подтверждением информации мтДНК: они указывают, что общий предок всех людей современного типа жил в Африке, а более поздний предок всех неафриканских народов — в Азии.
Наиболее серьезной проблемой для европоцентрической концепции культурного развития, в основе которой лежит гипотеза о северном маршруте исхода из Африки, является сам факт существования австралийских аборигенов, создавших свою собственную культуру пения, танца и живописи задолго до европейцев.
Каким образом и почему они попали в Австралию гораздо раньше, чем предки европейцев — в Европу? Эта загадка породила целый ряд попыток их объяснения.
В качестве доказательства присутствия еще в глубокой древности разумного человека на Леванте, в более узком смысле — Сирии, Израиля и Ливана, британский зоолог Джонатан Кингдон (Jonathan Kingdon, род. 1935) ссылается на многочисленные орудия эпохи Среднего палеолита, найденные в Индии. Возраст некоторых из них достигает 1бЗ 000 лет. Однако, серьезной проблемой является полное отсутствие скелетных останков человека современного типа такой древности где бы то ни было за пределами Африки.
Кингдон утверждает, что эти орудия могли быть изготовлены людьми предсовременного или архаического типа, которые как раз в то самое время обитали в Восточной Азии, и что древнейшая ветвь предков европейцев возникла и сформировалась где-то неподалеку от Индии.
Крайне маловероятно, что Северная Африка могла послужить источником генетического материала для жителей Азии. Все сказанное является аргументом в поддержку точки зрения о том, что Европа и Северная Африка являлись реципиентами (приемниками) миграций с Востока.
Генетическое древо сообщает нам о предках европейцев и жителей Леванта нечто весьма любопытное и неожиданное: оказывается, они пришли на эти земли не прямо из Африки, а откуда-то с юга, из районов, граничащих с Индией.
Почему некоторые археологи указывают на целый ряд культурных влияний на Европу в период между 20 000 — 50 000 лет назад, одно из которых исходило с Востока?
Существуют особые генетические маркеры, прослеживаемые параллельно двум различным волнам культурных влияний в археологически документированной истории Европы, которые имели место около 25 000 лет назад, накануне последнего ледникового максимума. Они показывают, что «восточные» корни европейцев — это отнюдь не нелепый вымысел.
Так, в Индии. присутствует ветвь мтДНК гаплогруппа U2i. Общий возраст этого клана на Ближнем Востоке превышает 50 000 лет. Согласно молекулярным часам, U5 также имеет возраст около 50 тысяч лет и представляет собой наиболее древнюю линию во всей Европе, возникшую за 15 000 лет до появления в Европе следующей линии-родоначальницы.
Но каким путем генетический маркер 54 000 — 50 000 лет назад распространился на Ближний Восток, а оттуда через дочернюю ветвь U5 в Европу, если археологические датировки появления наиболее ранних памятников людей Верхнего палеолита на Леванте и древнейших артефактов Ауриньякской (Ориньякской) культуры в Болгарии составляют соответственно 47 000 и 46 000 лет?

Ауриньякская (Ориньякская) культура (микролитические орудия труда и палеолитические «венеры») — археологическая культура раннего этапа позднего палеолита.
Это несоответствие можно объяснить систематическим занижением данных при радиоуглеродном анализе любого времени свыше 40 тысяч лет вследствие так называемого потолочного эффекта. Прародиной этих линий предков европейцев была Южная Азия. Ветвь U2i, на долю которой приходится около 9,5% всех материнских линий в Индии и свыше 78% всех линий — выходцев из Индии в Европе, несомненно, является коренной уроженкой этого региона. В Индии ее возраст составляет 53 000 лет.
Другим индикатором, указывающим на южноазиатский регион как на возможный источник генетических предков этого клана Европы, явилось нахождение там древнейших корней европейского типа и зарождение другой ветви, представленной в Европе, — гаплогруппы U7, которая впоследствии проникла на Ближний Восток и в Европу.
Три наиболее распространенные в Индии ветви мтДНК не встречаются больше практически нигде в мире (единственное исключение — некоторые районы Центральной Азии к северу от Индии) и, таким образом, словно эхо отражают это пышное цветение материнских линий в Индии. Две из них представляют собой линии потомков группы К, тогда как третья — это ветвь первого поколения, встречающаяся не только в упомянутых районах Центральной Азии, но и, что особенно важно, далеко на юго-востоке, в Камбодже. Все эти типы активно представлены у племен и этнических групп Южной Индии, особенно — у австралоидных ченчу. Люди племени койа, кроме того, сохранили и редкий корневой тип, ограниченный Индийским субконтинентом и соседними районами.
Наше материнское генетическое древо свидетельствует о том, что возраст древнейших выходцев из Южной Азии, ставших родоначальниками современных европейцев, составляет не менее 50 000 лет.
Другая линия потомков азиатов, также проникшая в Европу, характеризуется мутацией по мужской линии M17 (устаревшее название гаплогруппы R1a1 — снип М17 — прим. П. Олексенко). Встречаемость этой линии наиболее высока среди венгров, у которых она достигает 60%. Линия М17 доминирует во всей Восточной Европе, и хотя этот клан распространен практически во всех районах Западной и Восточной Европы, а также на Леванте, его встречаемость там остается стабильно низкой. Его минимальное присутствие на Леванте заставляет отвергнуть версию о том, что этот регион мог служить для М17 воротами на пути в Европу.

Питер Андерхилл (Peter A. Underhill), американский генетик https://news.stanford.edu/
Генетики Питер Андерхилл (Peter A. Underhill), Льюи Кинтана Мюрси (Lluis Quintana-Murci, род 1970, франко-испанский биолог и популяционный генетик, известный своими исследованиями эволюции человека, геномного разнообразия населения) и их коллеги в качестве наиболее вероятного кандидата на роль прародины линии M17, проникшей в Европу, считают Центральную Азию и особенно Алтай.

Тоомас Кивисилд (Toomas Kivisild, род. 1969), эстонский генетик
По мнению Оппенгеймера, а также по мнению эстонского популяционного генетика Тоомаса Кивисилда более логично предположить, что древнейшей прародиной линии M17 и ее предков была все же Южная Азия. Ведь именно там, в Пакистане, Индии и Восточном Иране и в меньшей степени — на Кавказе, мы находим наибольшее разнообразие вариаций M17.
Характерно, что M17 в Южной Азии отличается не просто гораздо более широкой вариативностью, чем в Центральной Азии; это многообразие вариаций характеризует ее присутствие в изолированных племенных группах Южной Азии, опровергая тем самым гипотезу о том, что M17 является маркером «проникновения мужского клана арьев» в Индию. Согласно одной из датировок, эта линия появилась в Индии около 51 000 лет назад.
Все это свидетельствует о том, что М17 первоначально могла совершить исход из Индостана через Кашмир, затем проникнуть в Центральную Азию и азиатскую Россию и, наконец, мигрировать на запад, в Европу.

Орнелла Семино (Ornella Semino) https://scholar.google.ru/
Поскольку происхождение европейской линии М17 к востоку от Европы, на Алтае, практически не вызывает сомнений, остается вопрос о датировке подобного исхода. Итальянский генетик Орнелла Семино (Ornella Semino) и ее коллеги, отказавшись от генетических данных, обратились к изучению линии М17 и ее «отца», M173 (гаплогруппа R1), и, рассматривая эти линии как составную часть общей палеолитической миграции с востока на запад, поддающейся археологической датировке, высказали предположение, что она имела место 30 000 лет назад.
Итак, мы упомянули M173 — еще одного серьезного кандидата на роль мужской линии, проникшей в Европу. Эта линия относится к очень обширному клану. Этот клан гаплогруппы R встречается повсюду — от Англии до Южной и Центральной Азии, в Сибири и даже дальше, вплоть до Австралии и Америки.
Немало постранствовал по свету, и, как говорится в одном исследовании, его коренной тип является самым распространенным типом одинарной Y-хромосомы в мире. Тем не менее, ареал расселения генетического «отца» — клана R ограничен Индией, Пакистаном, Центральной Азией и Северной Америкой.
Исследование географического расселения и многообразия генетических ветвей опять-таки показывает, что клан R вместе со своим «сыном», кланом M17, сформировался достаточно рано в Южной Азии, где-то в районе Индии, и впоследствии расселился не только на юго-восток, в Австралию, но и на север, непосредственно в Центральную Азию, где и разделился на две ветви, одна из которых устремилась на запад, в Европу, а другая двинулась в Восточную Азию.
В отличие от своего сына, клана M17, клан R не остановился на пороге Восточной Европы. Правильнее будет сказать, что на западе Европы его встречаемость очень высока, достигая 86% в Стране Басков и почти столь же высокого уровня — на Британских островах.
Это расселение «отца» и «сына» в разных направлениях свидетельствует о том, что первый мог проникнуть в Европу гораздо раньше последнего. Это можно объяснить влиянием Ледникового периода. А пока ограничимся тем, что скажем, что, хотя М17 в Европе можно считать сравнительным «новичком», его «отец» род R, по-видимому, является наиболее мощной мужской линией-маркером древнейшего проникновения в Европу с востока еще в эпоху Раннего Верхнего палеолита, около 30 000 лет тому назад.
Итак, на долю этой команды, «отца» и «сына», приходится примерно 50% всех существующих мужских генеалогических линий современной Европы.
Дополнительными аргументами в поддержку этой гипотезы являются результаты недавнего исследования Т. Кивисилда и его коллег. M17 имеет в Индии среднюю встречаемость порядка 27%, но достигает в одном из ее штатов, штате Пенджаб, уровня 47%. Наибольшего многообразия — куда более широкого, чем в Центральной Азии или Европе, — М17 достигает в Иране, Индии и Пакистане.
Это относится и к Южной Азии, которая является его древней прародиной. Необходимо отметить, что коалесцирующее определение для возраста M17, М173 и M172 по М89 (гаплотип Андерхилла 71) в Индии может составлять 88 300 лет — датировки по филогенетическому анализу (Kivisild, 2003).
Ключевой вопрос здесь — поиск М17 при высокой встречаемости и широком многообразии у представителей племенных групп Южной Индии, в частности у австралоидных племен ченчу и ядхава. Если бы M17 у представителей этих племенных групп была привнесенной, а не коренной линией, ее многообразие было бы значительно более низким.
На юге Индийского субконтинента основной тип внешнего облика меняется, и преобладающим становится так называемый дравидский тип, который дает немало оснований утверждать, что древней прародиной генетической линии гаплогруппы М является Индия. Дело в том, что именно там, в Индии, она имеет наибольшее многообразие и древность. Более нигде в мире она не обладает таким множеством вариаций и обилием основных ветвей.
Возраст древнейшей из ее «дочерей» в Индии, ветви М2, составляет около 73 тысяч лет. И хотя эта датировка времени распространения М2 не слишком точна, она вполне может отражать местное восстановление населения после гибели в результате катастрофического извержения вулкана Тоба около 74 000 лет тому назад. Ветвь М2 широко представлена у ченчу, народа охотников и собирателей, принадлежащего к австралоидной группе и живущего ныне в штате Андхра-Прадеш.
Ченчу обладают целым рядом уникальных местных вариантов ветви М2, а также имеют общего предка с типами М2, выявленными в других районах Индии. В целом есть все основания, чтобы считать местом возникновения клана М именно Индию, а не какой-либо другой район, находящийся к западу от нее или в Африке.
Самой же удивительной и любопытной среди всех родичей клана N в Индии является ее «дочь» гаплогруппа R, ставшая праматерью большинства людей Запада, включая Европу, не говоря уже о двух ее «дочерях» на Дальнем Востоке, каждая из которых тоже обладает обширным и разветвленным потомством.
При анализе присутствия линии R в Индии создается впечатление, что ее плодовитость напоминает планирование семьи. Эстонские генетики сумели идентифицировать многочисленные ветви потомков клана R, берущие начало в Индии, причем ни одна из них не встречается ни в каком другом регионе. Эти новые ветви настолько богаты, что удалось даже датировать время их распространения — около 73 000 лет тому назад.
Нигде более, ни на западе, ни на востоке, мы не находим столь широко разветвленного многообразия генетической линии R. Одного этого вполне достаточно, чтобы считать Южную Азию исходной прародиной клана R. Но, пожалуй, самое интересное здесь заключается в том, что невиданно пышный расцвет линий кланов М и N в Индии говорит о том, что они увековечили генетическую память о людях, которым посчастливилось выжить во время катастрофического извержения вулкана Тоба около 74 000 лет тому назад.
В той же Южной Азии существует целый ряд других туземных групп, включая так называемые прото-австралоидные этнические группы, такие, как корава, йанади, ирула, гадаба и ченьчу в Индии, а также ведды в Шри-Ланке. Недавнее исследование, посвященное двум из этих групп, ченьчу и койя, позволило выявить не только тот факт, что мтДНК и Y-хромосомные наборы их предков являются уникальными среди других народов Южной и Западной Азии, но и то, что они характерны для древнейшего генетического наследия всего этого региона.
По всей видимости, их предки-собиратели, образно говоря, заложили генетические семена для последующей дифференциации генофондов Западной и Восточной Евразии, и с тех пор представители этих народов испытали крайне ограниченный приток генов извне.
Андаманские острова
На Андаманских островах, расположенных в Бенгальском заливе, между Индией и Юго-Восточной Азией, живет целый ряд таких туземных групп, сохранивших архаические дописьменные культуры и обладающих резко выраженными чертами внешности так называемого африканского типа. К числу групп, менее всего затронутых внешним влиянием, относятся народности джарава и онге, представители которых живут на большинстве южных островов Андаманского архипелага. Результаты недавних генетических исследований, находящиеся в процессе анализа и изучения, способны помочь выяснить происхождение жителей этих островов как по материнской, так и по отцовской линиям.
Пути миграции древних южноазиатских народов
Наше путешествие во времени в глубины генетической и археологической истории человечества позволяет сделать два важнейших вывода: во-первых, генетической прародиной европейцев первоначально, около 50 000 лет тому назад, была Южная Азия, точнее — северо-запад Индостана и регион Персидского залива; и во-вторых, предки современных европейцев проникли в Европу, этот прохладный, но богатый сад, по двум совершенно разным маршрутам.
Более ранний из них пролегал через Плодородный Полумесяц, который около 51 000 лет тому назад был открыт в качестве своего рода коридора, ведущего от побережья Персидского залива. Коридор этот позволил мигрантам проникнуть в Турцию, Болгарию и далее, в Южную Европу. Это вполне совпадает с общим проникновением представителей Ауриньякской культуры в Европу.
Второй по времени маршрут из Южной Азии в Европу мог пролегать вверх по течению Инда, в Кашмир, и далее, в Центральную Азию, где примерно около 40 000 лет тому назад охотники начали охотиться даже на таких огромных животных, как мамонты. Некоторые из этих охотников, обладавшие передовыми по тем временам техническими навыками и орудиями, могли мигрировать на запад, за Урал, в европейскую Россию, а затем — на территорию современных Чехии и Германии.
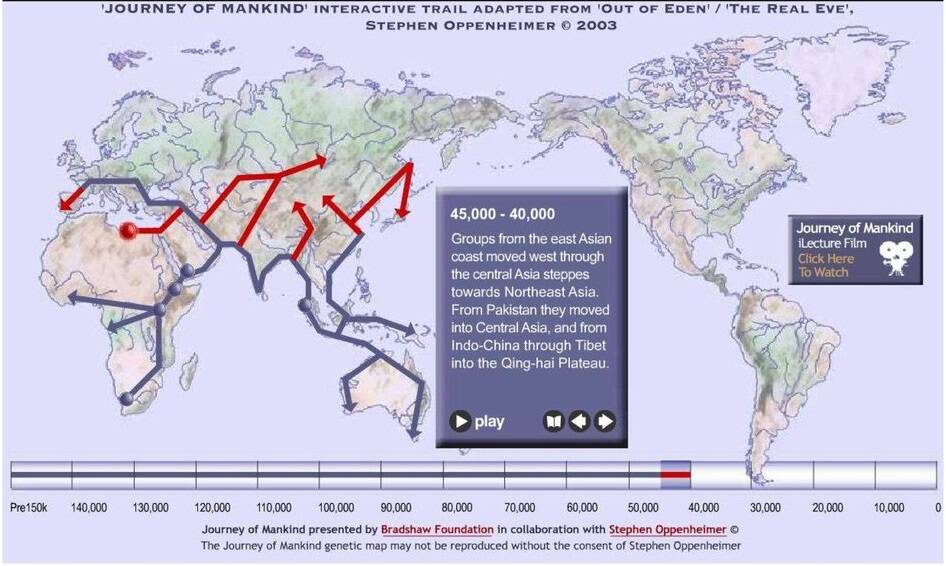
Древние миграции по Оппенгеймеру. Центрально-аазиатские и восточно-азиатсские миграации 45000 — 40000 лет назад https://bradshawfoundation.com/
Более консервативная точка зрения на это вторжение с востока сводится к тому, что наиболее ранним маршрутом проникновения человека современного типа на территорию России был путь через Закавказье, а не через Центральную Азию.
Составленное Генетическое древо говорит нам: это переселение отнюдь не было ранним исходом из Африки, как то полагают некоторые археологи и палеонтологи. Это было продвижение передовых групп на пути заселения всего Старого Света.
Индийский субконтинент, где отмечается широкое многообразие типов внешнего облика и культур населяющих его народов, представляет собой богатейшую этническую и географическую «транзитную зону» между Западной и Восточной Евразией. К северу и западу от нее, по мере продвижения из Индии в Пакистан, из Пакистана в Афганистан и Иран и далее в Европу, мы наблюдаем плавные и постепенные изменения внешнего облика коренных жителей, включая и пигментацию (цвет) кожи.
Внешнее сходство между европейцами и многими народами Южной Азии просто поразительно. Этот процесс постепенного изменения физического облика у жителей Северной Индии и Пакистана являет собой как бы параллель относительной частоте встречаемости двух генетических дочерних суперкланов.
Таким образом, северо-восточная часть Индийского субконтинента демонстрирует наиболее четкую и резко выраженную границу между востоком и западом. Эта граница, вполне возможно, отражает глубокую генетическую борозду, проходящую через всю Индию и связанную с облаком вулканического пепла, накрывшим Индийский субконтинент в результате извержения вулкана Тоба около 74 тысяч лет назад.
Древнейшие обитатели прибрежной полосы часто делились на все новые и новые ветви в различных пунктах своего долгого пути вдоль побережья Индийского и Тихого океанов и примерно 40 000 лет назад они заселили большую часть Азии и островов Восточного полушария. Первая из ветвей, двинувшаяся из Индии на север, стала родоначальницей охотников эпохи Верхнего палеолита, обитавших в степях Центральной Азии.
Последующие же ветви, расселявшиеся вдоль русел великих рек Центральной Азии, дали начало той самой расе, представителей которой мы сегодня называем монголоидами. Свидетельствами о существовании в Азии так называемого монголоидного типа вплоть до максимума Ледникового периода, имевшего место 20 тысяч лет назад, мы не располагаем.
 Прародина так называемого монголоидного типа внешности, если вообще можно говорить о каком-то едином комплексе признаков, остается неизвестной и спорной, хотя большинство имеющихся дентологических данных указывают на южные регионы. Вероятно, где-то в Центральной Азии (скорее всего — к северу от Гималаев) существовал особый регион, где в результате миграций, вызванных резким похолоданием и ухудшением климата, у коренных жителей Юго-Восточной Азии начали формироваться характерно «монголоидные» черты внешнего облика.
Прародина так называемого монголоидного типа внешности, если вообще можно говорить о каком-то едином комплексе признаков, остается неизвестной и спорной, хотя большинство имеющихся дентологических данных указывают на южные регионы. Вероятно, где-то в Центральной Азии (скорее всего — к северу от Гималаев) существовал особый регион, где в результате миграций, вызванных резким похолоданием и ухудшением климата, у коренных жителей Юго-Восточной Азии начали формироваться характерно «монголоидные» черты внешнего облика.
Итак, Передняя и Центральная Азия были заселены как бы тремя волнами генетической колонизации, берущими свое начало из Индии. Древнейшие поселенцы следовали по пути собирателей на прибрежной полосе, продвигаясь вдоль побережья из Индии через Индокитай к берегам Японии и Кореи, основывая на пути своей миграции колонии и поселения. Выходцы из этих поселений на побережье впоследствии проникли в глубинные районы Центральной Азии и вышли к берегам великих рек Азии, миновав проходы и перевалы в сплошной стене огромной горной гряды, протянувшейся с востока на запад и как бы обрамляющей Гималаи.
Первопроходцы продолжали продвижение в Юго-Восточную Азию и Австралию вдоль побережья Индийского океана. Они прибыли в Австралию примерно 60 тысяч лет назад, задолго до того, как началось активное заселение Европы.
Взглянув на генетическое древо, нетрудно понять, что, хотя современные австралийцы имеют с европейцами двух общих неафриканских предков по линиям М и N, живших около 70 тысяч лет тому назад, они тем не менее сохранили свои собственные характерные типы М и N. У нас нет никаких свидетельств того, что они, австралийцы, являются потомками европейцев. Нет у нас и никаких сведений о массовом «завозе» генов из Европы в Австралию в эпоху палеолита.
Европа и Передняя Азия были заселены значительно позже, около 50 000 лет тому назад, когда в мире наступило глобальное потепление, открывшее сухопутный коридор от района побережья Персидского залива к землям нынешнего Восточного Средиземноморья.
Археологические данные свидетельствуют о том, что Центральная Азия была заселена практически в то же время, что и Западная Евразия, то есть около 40 000 — 50 000 лет тому назад. Целая группа генетических линий, и сегодня присутствующих в Северной и Центральной Азии, подтверждают эту гипотезу и свидетельствуют о прямом пути продвижения на север с территории Индийского субконтинента до западных склонов Гималаев. Некоторые из этих северо- и центральноазиатских линий, по всей видимости, восходят к генофонду древнейших собирателей на прибрежной полосе в Юго-Восточной Азии, которые предположительно могли проделать долгий окружной путь и обойти Гималаи с запада.
См. еще:
Следопыт. Азиатская теория происхождения человека. Забытая реальность
Петр Олексенко, Игорь Токарев. Предки индийцев и других азиатов пришли из Африки?
Петр Олексенко. Индостан – колыбель человечества. Генетические исследования подтверждают древний возраст людей и обновление человечества 75 тысяч лет назад
Токар. Офтальмогеометрические расчеты Эрнста Мулдашева подтверждаются генетическими исследованиями
Leave a reply
Для отправки комментария вам необходимо авторизоваться.