Надежда Маркина. Археогенетика. Исследования Тоомаса Кивисилда и Y-хромосомная история Евразии и Америки
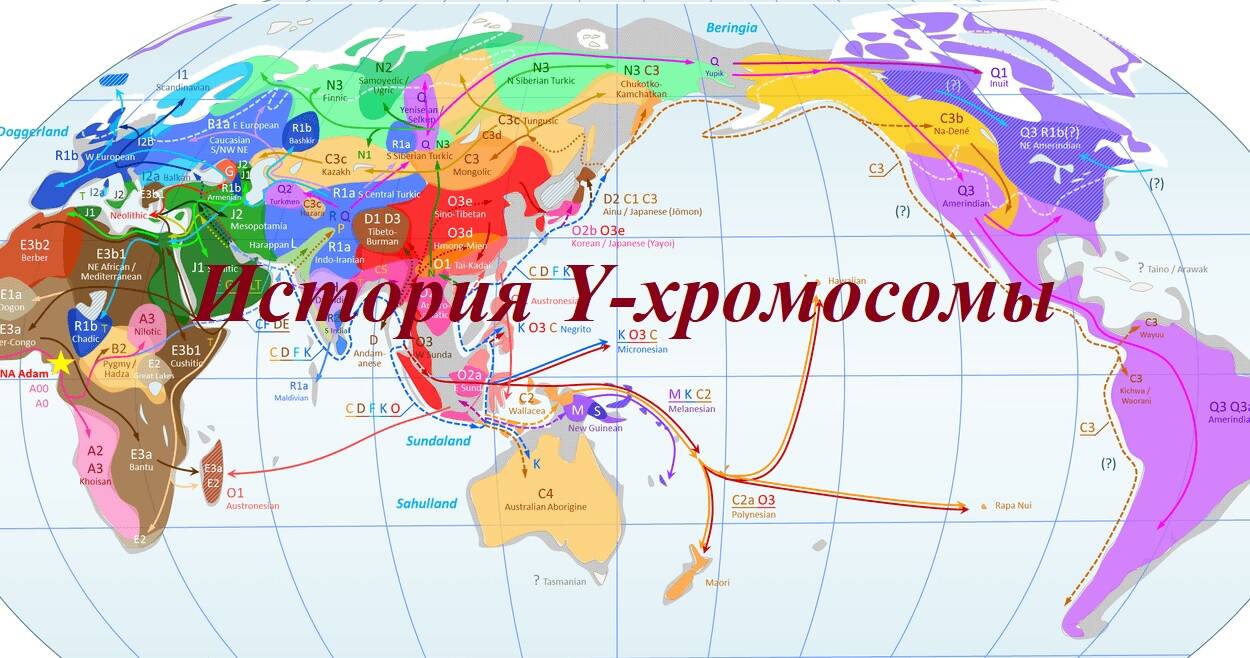
Предисловие П.Олексенко: древняя ДНК дает нам возможность напрямую исследовать, какие однонуклеотидные полиморфизмы (SNP) и гаплотипы Y-хромосомы присутствовали в разные периоды времени в регионах, поддерживающих долгосрочное выживание древней ДНК, и тем самым понять историю древних популций.
До недавнего времени исследования древней ДНК человеческих останков были сосредоточены в основном на вариациях, встроенных в митохондриальную ДНК (мтДНК). В этом обзоре рассматриваются свидетельства в масштабе генома о разнообразии древней Y-хромосомы, которые в последнее время начали накапливаться в географических районах, благоприятных для сохранения ДНК. Анализ последовательностей древней Y-хромосомы является сложной задачей не только из-за проблем, обычно связанных с работой с древней ДНК, но и из-за специфических свойств Y-хромосомы.
Древние последовательности Y-хромосомы дают нам первое захватывающее представление о древнем геноме и возможность оценить модели, основанные на ранее сделанных выводах из моделей генетической изменчивости в живых популяциях.
Автор перевода научного исследования — Маркина Надежда Вячеславовна, кандидат биологических наук.
* * * * *

Эстонский генетик Тоомас Кивисилд
Обобщение результатов, полученных при исследованиях Y-хромосом из древних геномов, от Тоомаса Кивисилда, одного из ведущих геномных специалистов, опубликовано в журнале Human genetics в марте 2017 года.
Профессор Тоомас Кивисилд, один из ведущих геномных специалистов, представляющий Кембриджский университет и Эстонский биоцентр, опубликовал обзор по исследованиям Y-хромосомы из древних геномов. В этой обобщающей работе он сфокусировался на данных по Y-хромосомному разнообразию древних популяций в разных регионах Северной Евразии и Америки.
Археогенетика на службе популяционных генетиков
Начало статьи посвящено методическим проблемам. На рисунке схематически представлены ошибки, которые могут возникать при исследовании древних образцов. Во-первых, чаще всего древнюю ДНК удается секвенировать с низким покрытием (не все нуклеотиды оказываются прочитаны даже по одному разу). Во-вторых, Кивисилд отмечает трудность различить истинные (прижизненные) мутации и мутации, происходящие из-за повреждений ДНК при длительном пребывании образца в окружающей среде, в особенности это касается замен нуклеотидов: C на T и G на A.
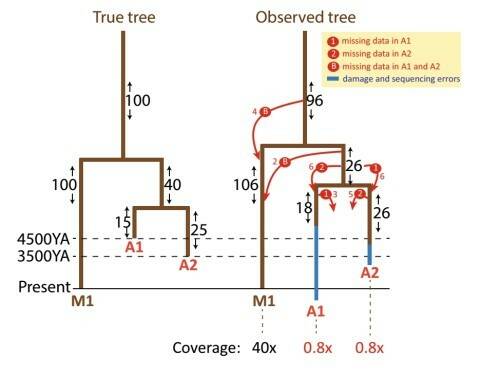
Рис. 1. Влияние низкого покрытия и post-mortem повреждений на длину ветвей и на картирование мутаций на дереве. Представлен пример филогенетического дерева, состоящего из одного современного образца, секвенированного с высоким покрытием (М1) и двух древних образцов, секвенированных с низким покрытием (А1 и А2). На обоих деревьях цифрами показано число мутаций между развилками дерева. Красными стрелками обозначены ошибки в числе мутаций.
Ошибки могут быть двух видов: пропущенные реально присутствующие мутации (из-за низкого покрытия) и обнаруженные не существующие в действительности мутации (из-за повреждений произошедших в ДНК за тысячелетия после смерти ее носителя).
Ошибки первого типа (ложноотрицательные). Из-за низкого покрытия часть реально присутствующих мутаций не обнаружена в одном или обоих древних образцах.
Если мутация пропущена в одном из древних образцов (стрелки 1 и 2), то на итоговом дереве она появляется только на ветви, ведущей ко второму образцу. Если мутация пропущена в обоих древних образцах, она на дереве появляется на линии, ведущей к современному образцу (который прочитан без ошибок). Ошибки второго типа (ложноположительные). Если образец А1 несет больше мутаций повреждения чем А2 (из-за большего возраста), то ветвь, ведущая к образцу А2, будет выглядеть намного длиннее, чем в действительности, а ветвь ведущая к А2 – лишь немного длиннее, чем в действительности.
В методической части обзора Кивисилд рассматривает два подхода к секвенированию Y-хромосомы: shotgun и hybridization-capture based. По методу shotgun («метод дробовика») ДНК фрагментируется на мелкие участки, которые секвенируют, а полученные перекрывающиеся последовательности собирают в одно целое. Этот метод применяется при секвенировании современных геномов; его же используют при секвенировании древних геномов с хорошей сохранностью, в этом случае он обеспечивает прочтение с высоким покрытием.
Технология hybridization-capture based — это целевое секвенирование. При этом считываются участки, содержащие определенные мишени (SNP), и это позволяет увеличить величину покрытия этих мишеней в итоговых данных. Метод позволяет получить достоверные данные даже в случае плохо сохранившейся ДНК, но его недостаток в том, что он не позволяет обнаружить новые вариации в геноме.
Y-хромосомная история Евразии
В соответствии с общим высоким генетическим разнообразием в Африке, наибольшее число глубоких разделенных ветвей Y-хромосомы отмечается в африканских популяциях. На других континентах основное Y-хромосомное разнообразие ограничивается тремя кладами: D, C и F. Внутри них основное расхождение приурочено ко времени 60-40 тыс. лет назад, в соответствии с моделью выхода из Африки. Кроме того, обнаруживаемые в популяциях Ближнего Востока и Европы молодые субветви гаплогруппы Е, вероятно, отражают эпизоды генетических потоков из Африки в течение последних 25 тыс. лет.
Три основных евразийских ветви D, C и F разделились на примерно 40 субветвей, возраст которых более 30 тыс. лет. На ветвях этого дерева можно расположить древние образцы (на основе данных по секвенированию Y-хромосом). На рис 3 показано расположение девяти древних образцов эпохи палеолита.
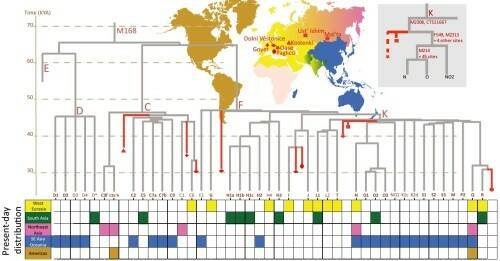
Рис.3. Разнообразие Y-хромосом за пределами Африки 20-50 тыс. лет назад. Разделение ствола на 41 ветвь (существующие Y-хромосомные линии) произошло не позднее 30 тыс. лет назад. Внизу разными цветами показано распределение субветвей в современных популяциях.
Два древнейших генома – Усть-Ишим (около 45 тыс. лет назад, на рисунке обозначен квадратиком) и Оase (37-42 тыс. лет назад, на рисунке обозначен звездочкой) оба расположены вблизи корня гаплогруппы К, относящейся к ветви F, которая объединяет большинство современных Y-хромосом. Оба этих образца находятся на ветви, предковой для гаплогрупп N и О, сегодня широко распространенных от финно-угорских народов до Тибета и Австралии.
Образец Костенки-14 (38-36 тыс. лет назад, на рисунке обозначен треугольником) принадлежит к гаплогруппе С, которая сегодня очень редка или отсутствует в большинстве европейских популяций, но часта в популяциях Сибири, Юго-Восточной Азии и Океании. Линия С присутствует также у иберийского охотника-собирателя (Ла Бранья) и у трех неолитических земледельцев из Анатолии и Центральной Европы. Это показывает, что линия С была широко распространена по Евразии до среднего голоцена, и впоследствии в генофонде произошли большие изменения. Распределение субветвей гаплогруппы С в древних и современных популяциях представлено на рис.4.
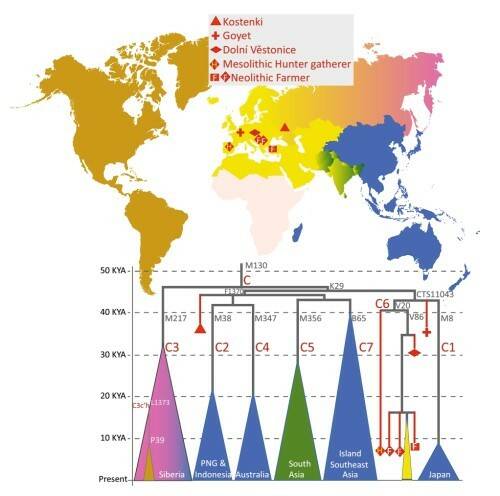
Рис.4. Основные субклады гаплогруппы С в древних и современных популяциях. Цвет треугольников внизу соответствует региону современного распределения субветвей, вершина треугольника указывает на время ветвления.
Помимо гаплогруппы С, Y-хромосомный пул охотников-собирателей и земледельцев раннего голоцена характеризуется линиями G, H, I, J и R.
Гаплогруппа G наиболее часта сегодня на Кавказе и в Передней Азии, но редка в Европе. Древние образцы из Анатолии и Ирана подтверждают, что G, наряду с Н, была наиболее обычной линией у ранних земледельцев в этих регионах. К этой же ветви принадлежит Y-хромосома «ледяного человека» Этци возрастом 5 тыс. лет назад (G2a-L91), она была обычной у анатолийских земледельцев, а сегодня наиболее часта на Сардинии и на Корсике.
Распределение гаплогруппы Н сегодня в основном ограничено Южной Азией, только одна субветвь H4-L285 встречается, причем редко, в Европе. Эта же субветвь была найдена у земледельцев Анатолии и Леванта, а также в образцах иберийского халколита.
Вероятно, линии G и Н были принесены в Европу с миграцией анатолийских земледельцев, пишет автор. Их частота остается высокой в некоторых ограниченных регионах, таких как Кавказ, Сардиния, Корсика, а в других частях Европы она падает. Распределение гаплогрупп G и Н в древних и современных популяциях представлено на рис.5.
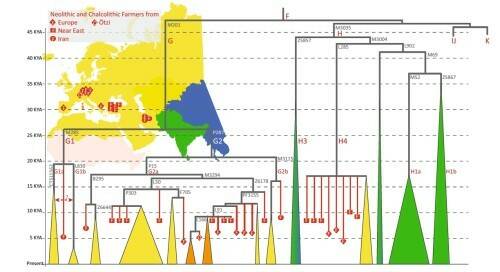
Рис. 5. Основные субклады гаплогрупп G и H в древних и современных популяциях. Цвет треугольников внизу соответствует региону современного распределения субветвей, вершина треугольника указывает на время ветвления.
География гаплогруппы I сегодня ограничена Европой. В древних образцах она обнаружена как у скандинавских охотников-собирателей, так и у анатолийских земледельцев и у земледельцев Центральной Европы. Ее наличие у современных европейцев может указывать на генетическую преемственность.
Гаплогруппу J связывают с распространением сельского хозяйства в Европу, но при этом ее не находят в европейском неолите, но находят у охотников-собирателей географически разных регионов, от Кавказа до Карелии, а также у двух земледельцев – из Ирана и из Анатолии. В Центральной и Западной Европе J начинает появляться в бронзовом веке, вероятно как результат движения популяций с Северного Кавказа. Распределение гаплогрупп I и J в древних и современных образцах представлено на рис. 6.
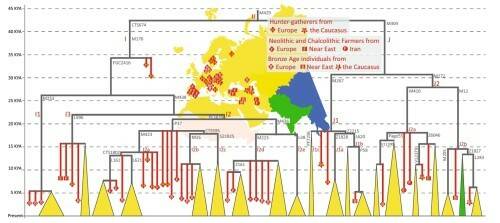
Рис. 6. Основные субклады гаплогрупп I и J в древних и современных популяциях. Цвет треугольников внизу соответствует региону современного распределения субветвей, вершина треугольника указывает на время ветвления.
Гаплогруппа R1b-M343 сегодня наиболее часта в Западной Европе, 90% достигает у басков, которые являются потомками донеолитической европейской популяции. Градиент снижения ее частоты с северо-запада к юго-востоку интерпретируется как результат смешения мезолитических охотников-собирателей с неолитическими земледельцами, принесшими гаплогруппы J и G.
Данные секвенирования показали, что время расхождения большинства ветвей R1b – 5-7 тыс. лет назад. К линиям R1a-M420 и R1b-M343 принадлежала большая доля Y-хромосом бронзового века в Центральной Европе, на Кавказе и в степном поясе Евразии. По мнению Кивисилда, большая часть современных линий R1a и R1b в Западной Евразии произошли от мужчин-основателей популяций позднего неолита – ранней бронзы. Распределение гаплогрупп R1a и R1b в древних и современных образцах представлено на Рис. 7.
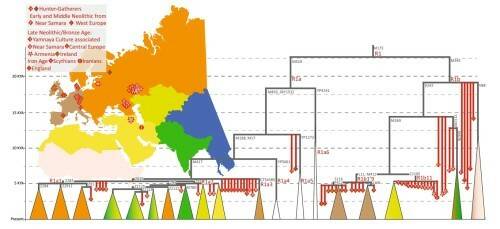
Рис. 7. Основные субклады гаплогрупп R1a и R1b в древних и современных популяциях. Цвет треугольников внизу соответствует региону современного распределения субветвей, вершина треугольника указывает на время ветвления.
Y-хромосомная история Америки
Современный генофонд американских индейцев – это смесь доколумбовой миграции из Сибири и поздних генетических потоков из Европы и Африки. Их Y-хромосомное разнообразие ограничено двумя линиями внутри гаплогруппы Q и одной или двумя — внутри C3-M217. Поскольку линии Q и С распространены в Евразии в разных регионах, можно предположить, что они достигли Америки разными путями. Вероятно, Q, наиболее частая у индейцев, происходит из Центральной Сибири, а менее распространенная в Америке C3-M217 — из Северной Сибири.
Важную информацию о формировании генофонда американских индейцев дает геном индивида со стоянки Мальта. Он оказался сходным с геномами американских индейцев и европейцев, но не восточноазиатских популяций, это дает основания предположить, что американские индейцы получили примерно треть предкового вклада от популяции, к которой принадлежит Мальта, и примерно две трети – от другой популяции, генетически близкой к Восточной Азии. Y-хромосома мальчика со стоянки Мальта ближе к западноевразийской R линии, но не к восточноазиатским D, C или O; она представляет собой вымершую гаплогруппу, которая отделилась от линии R вскоре после ее разделения на Q и R.
К линии Q1 принадлежат древние образцы Кенневикского человека (8,6 тыс. лет назад) и Анцик (12,6 тыс. лет назад) (см. рис. 8), в то время как Y-хромосома образца Саккак (4 тыс. лет назад) из Гренландии принадлежит к линии Q2, которая отделилась от Q1 более 25 тыс. лет назад. Это дает основания для вывода, что популяция палеоэскимосов (к которой принадлежал Саккак) была генетически замещена популяцией неоэскимосов.
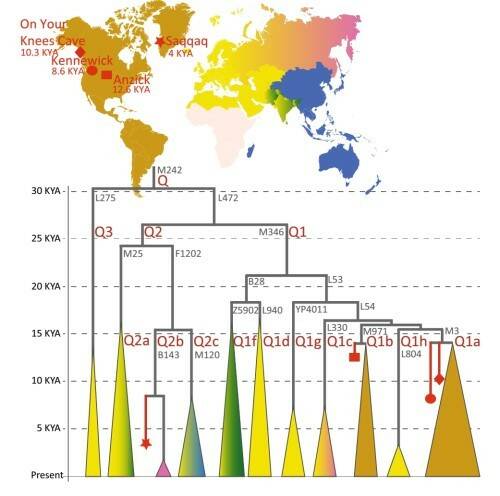
Рис. 8. Основные субклады гаплогруппы Q в древних и современных популяциях. Цвет треугольников внизу соответствует региону современного распределения субветвей, вершина треугольника указывает на время ветвления. Древние образцы обозначены красными символами (см. легенду вверху).
Подводя итог, Тоомас Кивисилд высоко оценивает возможности археогенетики, а именно секвенирования Y-хромосом из древних образцов для исследования генетического разнообразия в прошлом и реконструкции генетической истории человека. Есть надежда, что с дальнейшим увеличением числа древних геномов исследованиями будут охвачены больше географических регионов, в том числе регионы жаркого климатического пояса (пока что имеется только один успешно прочитанный геном из Эфиопии).
Источник: Генофонд.рф
См. еще:
Петр Олексенко. Индостан – колыбель человечества. Генетические исследования подтверждают древний возраст людей и обновление человечества 75 тысяч лет назад
Петр Олексенко, Игорь Токарев. Предки индийцев и других азиатов пришли из Африки?
Следопыт. Почему происхождение современного человека – пока еще загадка для науки


Leave a reply
Для отправки комментария вам необходимо авторизоваться.