Петр Олексенко. Индостан — колыбель человечества или перевалочный пункт в развитии цивилизаций?

О чем могут рассказать генетические исследования?
Сравнивая между собой гены разных людей в их ДНК из так называемой Y-хромосомы (которая передаётся непосредственно от отца к сыну без изменений из поколения в поколение), или из так называемой митохондриальной ДНК (она наследуется только от матери), исследователи находят в каких-то местах генетического кода различия в последовательности звеньев.
Чем меньше таких различий у представителей двух произвольных групп населения, тем в более близком родстве, видимо, находятся сами эти группы, тем недавнее они выделились из некоего общего «котла». А чем больше различий, тем отдалённее родство, тем протяжённее время раздельной жизни этих групп.
Используя средние скорости накопления таких генетических различий (а они приблизительно известны), можно определить, когда именно начали расходиться те или иные группы людей. Надо заметить, что ДНК митохондрий и Y-хромосом — единственные подходящие для таких исследований объекты, потому что первая вообще не меняется при делении клетки, а во второй есть участок, который при этом остаётся неизменным.
Генетическая гаплогруппа — это метка, возникающая в результате мутации генетической цепочки ДНК, которую можно проследить на протяжении многих поколений по мужской линии. Эта мутация обозначается определенным маркером. Гаплогруппа не имеет никакого отношения к цвету глаз, волос, форме черепа, умственным способностям, темпераменту. Но генетика с помощью этой метки может рассказать о путях миграции больших групп людей в те времена, от которых почти не осталось даже археологических свидетельств.
Например, азербайджанец может считать себя хоть тунгусом, это его право, но при этом он все равно будет азербайджанцем. А вот дети этого азербайджанца, при условии, что жена азербайджанца русская, могут быть на выбор — русскими или азербайджанцами. Внуки этого азербайджанца, при условии, что один из их родителей русский, вряд ли будут иметь выбор, они будут русскими не только по языку и культуре, но и по крови (на 75%). Это есть ассимиляция на культурном и генетическом уровнях.
Справедливости ради надо сказать, что внуки могут иметь 25% азербайджанской крови по мужской линии, если у азербайджанца родится сын. Y-хромосома сохранит азербайджанское происхождение. А если у него будет дочь, тогда его внуки будут иметь гаплотип, переданный русским отцом.
Если русский моряк, посетив Анголу, «подарит» туземке ребенка-мальчика, то у него, и у всех его потомков по мужской линии, будет гаплогруппа отца. Сменится 1000 поколений, потомки по всем параметрам превратятся в самых типичных ангольцев, но все равно будут носить европейскую Y-хромосому. И никаким другим образом, кроме анализа ДНК, это факт выявить не получится.
Мексиканцы — это латиноамериканцы, но это не означает, что латынь появилась в Мексике. А вот в Биробиджане очень большой процент еврейского населения, но это не означает, что Биробиджан — родина евреев.
В многочисленных исследованиях было показано, что генетическое разнообразие людей на 90% обусловлено различиями, возникавшими внутри популяций, живших на каком-нибудь континенте, и лишь на 10% — различиями между континентальными популяциями. Иными словами, в среднем генетические различия в пределах какой-либо популяции людей выражены гораздо сильнее, чем различия между двумя популяциями.
А это значит, что представители различных популяций отличаются друг от друга лишь чуть больше, чем представители одной и той же популяции.
Но представители одной и той же расы могут значительно отличаться друг от друга своими генами. Соответственно, у представителей разных рас может быть большее генетическое сходство, чем у представителей одной и той же расы.
Генетические исследования показали, что люди, которые на первый взгляд должны были произойти из одного континента и относиться к одной и той же расе, в реальности оказываются членами совсем разных групп. Эти группы и есть те древнейшие изолированные группы населения земли, которые долгое время развивались независимо друг от друга и начали смешиваться сравнительно недавно.
Генетические исследования в Индии
В 2004 г. в рамках проекта «Индийское генетическое разнообразие» на основе исследований ДНК у 1871 жителя Индостана из различных этнических, религиозных и племенных групп была составлена первая генетическая карта населения Индии. Так, геномы проживающих в Кашмире индуистов из высших каст оказались идентичными с проживающими там же мусульманами, и у них же много общего с генами представителей дравидийской группы. У дравидов, в свою очередь, ученые установили генетическую связь с «индоевропейцами».
А в 2009 г. самое масштабное исследование ДНК граждан Индии проводили исследователи из индийского Центра клеточной и молекулярной биологии (Centre for Cellular and Molecular Biology — CCMB), американской медицинской школы Гарварда (Harvard Medical School), школы здравоохранения того же университета (Harvard School of Public Health) и института Броуд в Бостоне (Broad Institute of Harvard and MIT).
Учёные проанализировали около 500 тысяч генетических маркеров (SNP), которые были вычленены из ДНК 132 человек из 25 различных групп со всех 13 штатов Индии. Были представлены все шесть языковых групп, а также различные касты и племенные группы (подробнее об этом исследовании в опубликованной статье в журнале Nature).
Довольно сенсационным оказался вывод генетиков о том, что все индийцы произошли от двух древних сильно отличавшихся друг от друга групп предков. Различные индийцы унаследовали от 40 до 80% генов от популяции, которую условно назвали Северные индийские предки (Ancestral North Indians — ANI). Они в свою очередь были чем-то похожи на жителей Ближнего Востока, Средней Азии и Европы. Вторая группа прародителей индийцев была условно названа Южные индийские предки (Ancestral South Indians — ASI), эти люди не были похожи ни на один народ, проживающий за пределами Индии.
По словам одного из авторов работы Дэвида Райха (David Reich), в ходе исследования выяснилось, что представителям высших каст, а также людям, говорящим на индоевропейских языках (хинди и др.), досталось больше генов «северных предков». У остальных преобладали южные корни, а наиболее «южными» оказались коренные жители Андаманских островов в Индийском океане.
По словам ведущего исследователя из CCMB Кумарасвами Тангараджана (Kumarasvami Thangarajan), заселение Андаманских островов и южной части Индии происходило 65 тысяч лет назад. Позднее, 40 тысяч лет назад, появились северные индийцы. В какой-то момент времени древние север и юг смешались, порождая разнообразный колорит генов индийского населения. Сразу за массовым смешением ANI и ASI последовал период долгой (тысячелетней) изоляции отдельных групп, в основном, благодаря разделению на касты.
Таким образом, генетические исследования показали, что жители Индии несут в себе гены как европейцев, так и азиатов. Индия оказалась на перекрестке генетическим «течений» из Европы и Азии. В связи с этим вызывает удивление факт, что в Индии, несмотря на большое количество чужих культур и языков, в целом присутствует собственный генофонд. Он не замещался и не вытеснялся никогда.
Некоторые специалисты также считают, что жители Индии и Пакистана в определенном смысле отличаются от своих соседей и могли бы сформировать отдельную седьмую расу, так называемую «индо-дравидскую расу», находящуюся где-то посередине между европейской и азиатской, но ближе к европейской.
Северные индийские предки
Давайте сначала посмотрим, что представляют собой «северные индийские предки» (Ancestral North Indians). Ведь с ними, в первую очередь, связывают понятия «арии» и «индоевропейцы». Сегодня многие специалисты считают, что распространение индоевропейской семьи или индоевропейских языков ассоциируется с гаплогруппой R1a1 Y-хромосом, которая отождествляется с ДНК-маркером M17.
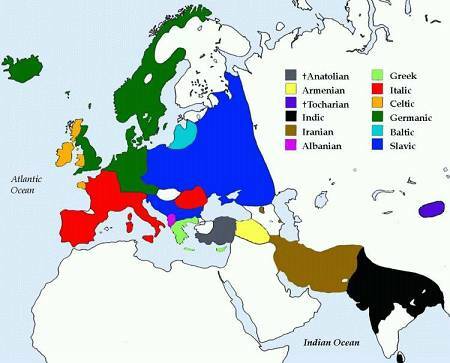
Карта распространения индоевропейских языков
Многие специалисты от традиционной науки, особенно историки, лингвисты, этнографы, в «арийском» вопросе исходят из банального европоцентризма и представления о цивилизаторской роли европейцев вообще и древних ариев как их представителей, в частности. Так уж сложилось, что проблема происхождения «индоевропейцев» очень тесно связана с политикой.
Понятно, что германский миф нам очень дорого обошелся. У «анатолийской версии» тоже есть свой политический подтекст. Версия о ностратической общности (некоем общем центре многих цивилизаций) также имеет ярых сторонников и ярых противников.
Сегодня наибольшая концентрация маркера М17 (около 40%) отмечается у мужчин, проживающих на всём протяжении от Чешской Республики, Балкан до степей Сибири и юга Центральной Азии. В Индии его имеют около 35% мужчин, говорящих на языке хинди, а в сообществах, говорящих на дравидийских языках — около 10% мужчин. Около 35%, мужчин, проживающих в восточной части современного Ирана, также имеют маркер М17. Он найден и у 5-10 % ближневосточных мужчин.
Вопрос о локализации прародины гаплогруппы R1a1 всё ещё остаётся спорным. Предположительно, она возникла в Евразии. На сегодняшний день есть два очага её наивысшей концентрации — в Северной Индии и на Украине, а также на юге России. С одной стороны, высокая частота встречаемости может наблюдаться в любом регионе с высокой численностью населения, с другой стороны — ее наивысшее разнообразие наблюдается на Балканах.
Предком R1a1 была гаплогруппа R1a, которая произошла от R1. Согласно современным представлениям гаплогруппа R Y-хромосомы человека (отмечена мутацией в SNP маркере M207) появилась около 30-35 тысяч лет назад предположительно на северо-западе Азии. Однако большинство разновидностей гаплогруппы R найдено среди населения Средней Азии, Сибири и Индийского субконтинента.
Поэтому, прародину гаплогруппы R, скорее всего, следует поместить туда, где есть наивысшее разнообразие её субкладов, то есть место, где рядом живут люди с R1*, R2*, R1a и R1b. Это — стык современных Китая, Афганистана, Северной Индии и Пакистана: Пенджаб — Памир — Синьцзянь.
Предок гаплогруппы R род P (M45) встречается в небольшом количестве в Индии, Пакистане, Китае, и Тибете. Предположительно он возник около 40 000 лет назад к северу от Гиндукуша, горной цепи в Афганистане и северных областях современного Пакистана. Потомки этого рода, носители гаплогруппы R (M207), мигрировали к северу от Гиндукуша в Казахстан, Узбекистан и южную Сибирь. От рода R появились R1 (отмеченная мутацией M173) и R2(M124).
Надёжных данных по возрасту R2 нет. Поэтому, трудно сказать, кто возник раньше — R1 или R2. Гаплогруппа R1 предположительно зародилась в Центральной или Южной Азии 25-30 тысяч лет назад, Её два основных субклада R1a (M420) и R1b (M343) (прочие варианты встречаются исключительно редко) являются наиболее распространёнными во всей Европе и Западной Евразии.
Гаплогруппа R2 возникла около 30 тысяч лет назад в Средней Азии. Позднее, примерно 25 тысяч лет назад она начала распространяться в Индии. R2 встречается (>15%) в верхних кастах у индусов, у буришей, чеченцев, курдов, парсов, сингалов, цыган и ограничена, в основном, территорией Индии, Ирана и Центральной Азии и Кавказа. Отдельные случаи обнаружения этой гаплогруппы отмечены среди населения Европы. За пределами этих регионов этот маркер полностью отсутствует.
Гаплогруппа R1a (М420) появилась 18 — 20 тысяч лет назад в Азии или Южной Азии. Это согласно исследованиям, прежде всего, американского генетика Питер Андрехилла и его коллег. Другие исследователи включают в ареал ее распространения также Центральную Азию, Ближний Восток и даже Восточная Европу. R1a имеет высокую частоту (>45%) у поляков, украинцев, белорусов, русских, казаков, пуштунов, таджиков, киргизов, алтайцев, марийцев. R1b (М343) имеет высокую частоту (>55%) у валлийцов, ирландцев, басков, португальцев, британцев, голландцев, итальянцев, фризов, бельгийцев, каталонцев.
Распространение гаплогрупп по языковым семьям
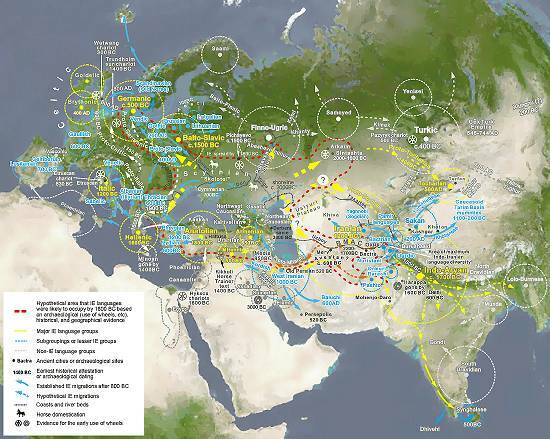
Карта распространения индоевропейских языков. Красная пунктирная линия — гипотетическая зона распространения индоевропейских языков к 1800 г. до н.э. (создана на основе археологических данных (использование колеса, приручение лошади и др.), исторических и географических событий. Желтая пунктирная линия — группы основных индоевропейских языков. Синяя пунктирная линия — подгруппы или малые индоевропейские группы. Светло-серая пунктирная линия — группы неиндоевропейских языков. Mohendjo-Daro — древние города или археологические поселения. 1400 г. до н.э. — самое раннее историческое упоминание или археологическая дата. Синяя стрелка — миграция индоевропейцев после 800г. до н.э. Синяя пунктир. стрелка — предполагаемая миграция индоевропейцев. Темно-серая пунктирнаая линия — границы и русла рек. Район одомашнивания лошади
» R1a
ИЕ: славянские, индо-иранские, балтийские;
Алтайские: тюркские;
Уральские: финно-угорские;
Западнокавказские
» R1b
ИЕ: кельтские, италийские, германские, армянские;
Алтайские: тюркские;
Восточнокавказские;
Изолянт: язык басков
» R2
ИЕ: индо-иранские;
Дравидские;
Восточнокавказские;
Изолянт:Б урушаски
«Индоевропейская» гаплогруппа R1a и вероятное место ее возникновения
Остановимся подробнее на гаплогруппе R1a. Совсем недавно именно с этой гаплогруппой связывали «индоевропейцев» и предполагаемое «арийское вторжение» в Индию. Ареал распространения R1a — от Исландии (викинги) до Индии (каста брахманов), современный центр высокой концентрации гаплогруппы находится на территории Польши — 56.4% .
R1a встречается у таджиков 64%, киргизов 63%, венгров 56%, украинцев 54% русских 47% , белорусов 47 %, казанских татар 34 %, македонцев15.2% , болгар 14.7%, герцеговинцев 12.1% . у народов скандинавского происхождения в Исландии 23 %, в Швеции и Норвегии 18-22 %, в восточном Иране 25 %.
Среди казахов — во всем остальном типичных монголоидов — носителей группы R1a не меньше, чем в России. Она также встречается в западных областях Китая и в Восточной Сибири. R1a присутствует в значительной степени в Пакистане, в южной Индии у дравидийских групп (до 26%) в штатах Андхра-Прадеш , Тамилнаду.
Индийские исследователи подтвердили различные присутствия R1a даже среди низших каст и индийских племен, не входящих в кастовую систему. Однако, гаплогруппу R1a в Y-хромосоме у мужчин нельзя считать маркером индоевропейского происхождения, так как по данным скорости мутаций Y-хромосом мутация R1a возникла более 10 тысяч лет назад, то есть намного раньше расселения прото-индоевропейцев.
Более того, недавние генетические исследования показали, что индийская гаплогруппа R1a имеет более разнообразное повторение в Индии и Иране по сравнению с Европой и Средней Азией, а это означает, что Южная и Передняя Азия могла бы быть источником ее возникновения.
Гаплогруппа R1b, наиболее распространённая во всей Европе и Западной Евразии, проявляет самое высокое разнообразие в районе Восточной Турции и Западного Ирана. Таким образом, R1b, скорее всего, возникла на иранском плато. Более того, R1b — также наиболее вероятный кандидат на то, чтобы быть «индоевропейской» гаплогруппой народов «белой расы».
Различная интерпретация времени и места появления гаплогруппы R1a1
Разные исследователи по-разному интерпретируют результаты генетических исследований, причем часто их оценки становятся диаметрально противоположными. Результаты оценки возраста напрямую зависят от применяемого метода, из-за чего пока не существует общепринятой точки зрения о времени и месте появления гаплогруппы R1a1.
А. Носители гаплогруппы R1a1 — древние арии и славяне
1. Интерпретация С. Уэллса из Национального географического общества (США)
Национальное географическое общество США (National Geographic Society) и компания IBM создали исследовательский проект (Genographic_Project), в рамках которого использован очень сложный лабораторный и компьютерный анализ ДНК сотен тысяч людей, включая туземцев и обычных жителей многих стран и городов, чтобы составить карту заселения Земли современными людьми.
Так, в рамках этого генографического проекта специалисты определили, что маркер M17 (гаплогруппа R1a1) появился в регионе современной Украины или южной России предположительно 4500 лет назад. Генетик и антрополог Спенсер Уэллс (Spencer_Wells), научный сотрудник Национального географического общества США, несколько лет возглавлявший проект «География гена», утверждает, что прародина ариев находится за пределами Индии (см. статью The Eurasian Heartland: A continental perspective on Y-chromosome diversity).
2. Интерпретация Антолия Клесова (США)
Карта Евразии
В своих изысканиях в области ДНК-генеалогии, российский исследователь, проживающий в Бостоне, доктор химических наук А. Клесов отождествляет носителей гаплогруппы R1a1 (маркер М17) с древними ариями и славянами. По его расчетам, возраст предка R1a1 (ариев) составляет 20 тысяч лет, а прародитель ариев жил в Южной Сибири. Отправной точкой миграции праариев стали Балканы 12 тысяч лет назад.
Далее они стали распространяться по Европе. 4900 лет — расчётный возраст предка ариев, положившего начало всем носителям, ныне живущим на Среднерусской равнине, 4000-3600 лет назад арии появились на Южном Урале, 3500 лет назад — в Индии; ими оставлен глубокий след на пути в Индию — до 64% таджиков и киргизов являются носителями R1a1.
(Более подробно о статьях А.Клесова см. Приложение № 1 Краткое эссе по публикациям А. Клесова)
Давайте нарисуем карту миграций «ариев» по Клесову. Получается, что предки ариев жили в Южной Сибири 20 тысяч лет назад, затем они, не оставляя после себя никаких «материальных» и «культурных» следов, пришли на Балканы 12 тысяч лет назад. С этого момента они стали протоиндоевропейцами. Прожив на Балканах почти 6000 лет, они со своим арийским языком начали расселяться по всей Европе и в восточном направлении по территории современной России.
4900 лет назад арии сформировались в этническую группу славян (место максимальной концентрации R1a1 — этнический очаг). Русский народ быстро расплодился и стал расширять ареал своего обитания. Поэтому R1a1 является, согласно Клесову, биологическим маркером русского этноса. Через 1000 лет славяноарии продвинулись далее на Урал, построив «цивилизацию городов» (Аркаим и др.) со множеством медных рудников и международными связями вплоть до Крита.
А еще через 500 лет они пришли через Памир в Индию, создали санскрит и написали Веды. После прихода в Индию 3500-3600 лет назад арии, носители гаплогруппы R1a1 стали «индоевропейцами».
Другая миграция славяноариев через южный поток 4200 лет назад прошла через Среднюю Азию. Прожив там 500-800 лет, арии перешли в Иран и написали там Авесту. Общие предки индийцев и иранцев гаплогруппы R1a1 жили около 4100 лет назад, то есть на 800 лет раньше общего предка современных этнических русских гаплогруппы R1a1.
3. Интерпретация Н. Максимова
Приведем несколько интересных фрагментов из статьи Н. Максимова «Геногеография — путь в наше прошлое«:
«Особая роль Балканского узла в истории Европы, как свидетельствует геногеография, определялась тысячелетия назад и определила собой историческую географию обоих европейских генофондов — восточного и западного…
Ну и поразительна встреча двух культур на Урале. Разные генофонды (в рамках генофонда Евразии) к западу и к востоку от Урала в первую очередь обусловлены тем, что к западу и к востоку от Урала обитают разные расы — европеоиды и монголоиды. Видно, что генетическая граница между Европой и Азией проходит там же, где и географическая — по Уралу.
Когда люди расселялись в разные стороны, мутации в географически изолированных популяциях накапливались, давая начало новым генетическим линиям. Но популяции не только разделялись, в какие-то периоды они смешивались…
Не так давно достоянием научной общественности стали результаты генетического исследования останков представителей древнего населения Сибири. Учеными были исследованы 25 костных фрагментов, относящихся к четырем древним культурам — андроновской, карасукской, тагарской и таштыкской, которые существовали на территории современного Красноярского края с третьего тысячелетия до н.э. по четвертый век н.э.
Восточный и западный генофонд
Цель исследования заключалась в том, чтобы по аутосомным генам определить цвет кожи, волос и глаз людей. Это удалось сделать для 23 из 25 образцов. Оказалось, что, по меньшей мере, 60% исследованных лиц имели светлые (голубые или зеленые) глаза.
Вывод ученых: «фенотип большинства древних образцов имел типичную светлую европейскую пигментацию, по всей видимости, им были присущи черты типичного европейского пигмента, т.е. глаза голубого или зеленого цвета и светлые волосы и кожа, и они, вероятно, имели индивидуальное европейское происхождение»….
В том же исследовании гены из 25 костных останков были сравнены с образцами генов европейцев, азиатов и африканцев. Оказалось, что 22 лица имели европейское происхождение, 2 — азиатское и 1 — промежуточное между европейским и азиатским. Ранее те же останки были исследованы с целью определения их мужской гаплогруппы (Y-хромосомы). Это удалось сделать для 18 образцов. 17 из них относились к индоевропейской гаплогруппе R1a1, а 1 — к азиатской гаплогруппе С.
Таким образом, очевидно прямое соотношение между индоевропейской мужской гаплогруппой R1a1, европейскими аутосомными генами и светлой пигментацией».
4. Интерпретация А. Тюрина
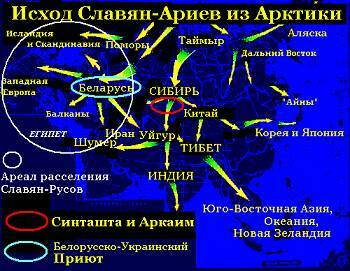
Карта Русь-Орды по Тюрину
Согласно еще одному российскому исследователю Анатолию Тюрину, генетическая Y-гаплогруппа R1а является элитной гаплогруппой именно славян — ариев.
В своей статье «Русь-Орда и генетические карты Старого Света» он пишет: «Три массива генетической информации — карты распределения групп крови, данные, обосновывающие наличие общего предка у славян и популяций Северного Индостана, а также характерное распределение европеоидности в кастах популяций Южного Индостана, прямо и однозначно голосуют за соответствие действительности одной из самых ярких гипотез Новой Хронологии А.Т. Фоменко и Г.В. Носовского — арии пришли в Индию из Руси-Орды».
В противоположность приведенным выше оценкам есть и другие мнения и расчеты.
Б. Носители гаплогруппы R1a1 — древние индийцы
5. Интерпретация Томаса Кивисилда (Эстония), Сенгупты (Индия), Сахо (Индия), С. Шармы (Индия)
Эстонский биолог Томас Кивисилд (Toomas Kivisild) показал в своем исследовании в 2003 г. максимальное разнообразие в R1a1 (M17/M198) на территории Южной Азии. Кивисилд считал, что источником гаплогруппы R1a1 была Южная и Передняя Азия. Это утверждают и индийские исследователи Сенгупта (Sengupta, 2005), Саха (Sahoo, 2006), С. Шарма (2009). Авторы рассчитали «разнообразие» гаплотипов гаплогруппы R1al, и нашли, что оно равно 0.52 в Индии, 0.40 у европейцев и 0.32 в Средней Азии.
Как отмечают индийские исследователи, вопрос происхождения гаплогруппы R1a1 остается не совсем ясным еще и потому, что обнаружено слишком мало носителей родительских гаплогрупп R*, R1* и R1a*, как и субкладов R1a1a, R1a1b и R1a1c.
6. Интерпретация П. Андерхилла и Ш. Мирабаль (США)
Питер Андерхилл (2009) , Regueiro (2006), показали, что R1a и R1a1 происходит из южно-азиатских стран. Так, согласно исследованиям американских генетиков Питера Андерхилла (Peter A. Underhill) и Шейлы Мирабаль (Sheyla Mirabal) из Департамента молекулярной и человеческой генетики и Международного университета в Майами (Флорида) в 2009 г., гаплогруппа R1a1 является теперь R1a1a.
Имеется два главных субклада — R1a1a* и R1a1a7. R1a1a7 с маркером M458 — европейский маркер, и его эпицентр — Польша. В Европе представлены субклады R1a1a* и R1a1a7 (M458), в Южной Азии — R1a1a*, R1a1a5 (PK5) и R1a1a6 (M434). Доля R1a1a7 в Европе — до 70% гаплогруппы R1a1 и ее носителями является более 30% населения в основных областях Восточной Европы.
По оценкам авторов, возраст восточно-европейского субклада R1a1a7 составляет 10700 лет, а общий возраст R1a1a* в Европе — 11300 лет. Общий возраст R1a1a* в Западной Индии — 15800 лет. В связи с этим авторы высказывают предположение о южно-азиатском происхождении данной гаплогруппы.
Андерхилл также утверждает, что никаких древних миграций из Восточной Европы в Индию не было, поскольку в Индии отсутствует восточно-европейский субклад R1a1a7. По его мнению, речь может идти лишь о древней миграции R1a1a из Азии в Европу, а субклад R1a1a7 возник в Восточной Европе уже после периода появления ариев в Индии.
Работы этих исследователей является самым свежими и всеобъемлющими. Они ставят под сомнение исход носителей R1a1 из Восточной Европы, о котором пишут другие исследователи.
Идем дальше. Иследования П. Андерхилла и Ш. Мирабаль показали, что по оценкам всех возрастных вариантов своего происхождения славяне сформировались уже после того, как R1a1 рассеялся настолько широко, что смог объединить Центральную Европу и Индию. Так что роль славян как распространителей гаплогруппы не объясняет происхождение и расселение R1a1 в целом. Чем больше данных, тем обоснованней это предположение.
Андерхилл и Мирабаль рассчитали время появления R1a * и R1a1 в Индии — соответственно 18 478 и 13 768 лет, то есть намного раньше, чем появление там индо-арийской языковой семьи. В исследовании высказывается предположение об автохтонности, т.е. местном происхождении северных индийцев и происхождении R1a1 на Индийском субконтиненте.
7. Интерпретация С. Оппенгеймера (Англия)
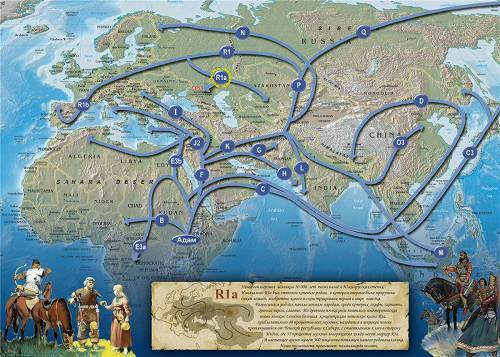
Миграции по Опенгеймеру
Английский исследователь Стивен Оппенгеймер (Stephen Oppenheimer) в своей книге «Изгнание из Эдема» (2004) написал: «Для меня и для Томаса Кивисилда, Южная Азия — родина маркера M17 и его предков. Мы находим самые высокие показатели и самое большое разнообразие линии M17 в Пакистане, Индии, и восточном Иране. Бол ее того, это разнообразие присутствует в изолированных племенных группах на юге Индии, таким образом, подрывая любую теорию M17 как маркера ‘мужского арийского вторжения’ в Индию.
По нашим расчетам, предполагаемый возраст происхождения этой линии в Индии — 51 000 лет. Все это предполагает, что M17, возможно, начал свой путь первоначально из Индии или Пакистана, через Кашмир, затем через Среднюю Азию в Россию и затем в Европу».
Есть сведения о том, что в ВОГИСе (Вавиловское Общество Генетиков и Селекционеров) был сделан доклад по теме генетики, на котором показали карты для субкладов R1a1f (или R1a1a7), R1a1f1, которые разделяют европейские и азиатские R1a1. Имеются совсем новые и неопубликованные снипы, которые пока никто кроме авторов не типировал. Они якобы хорошо делят R1a1 территориально, разделяя европейские и индийские маркеры.
Андерхилл и Хаммер владеют огромной коллекцией приватных снипов разных гаплогрупп, из которых опубликована только небольшая часть. Все секреты принадлежат им. Андерхилл нашел еще 5 дополнительных снипов для R1a1 в придачу к M17/M198 и новые субклады.
Вот дерево R1a1 по номенклатуре Андерхила: R1a1 M17, M198… новые M417, M512, M514, M515; R1a1a M56; R1a1b M157; R1a1c M64.2, M87, M204; R1a1d P98; R1a1e PK9; новые: R1a1f M458; R1a1f1 M334; R1a1g M434; R1a1h М?. По-видимому, именно здесь нам следует ждать разгадки о происхождении так называемых северных индийских предков или индоариев.
Давайте теперь посмотрим на некоторые нюансы времени и места возникновения «арийской» гаплогруппы R1a1. Возраст европейской, среднеазиатской и индийской гаплогруппы R1a1, определенный по STR, одного порядка: Пакистан — 12600 — 18600 лет (средн. 15600), Оман — 9600 -15400 лет (средн.12500), Западная Евразия — 10300 — 15300 лет (средн. 12500), Центральная Азия — 6200 — 16200 лет (средн.11200), Турция — 7400 — 12600 лет (средн. 10000), Греция — 6500 — 12100 лет (средн. 9300). У киргизов (63% R1a1) такие же древние R1a1, как в Индии и Европе.
R1a1 присутствует у многих, даже у современных корейцев, японцев (особенно у айнов на острове Хоккайдо), а также среди индейцев Аляски, были сообщения о том, что недавно нашли древние R1a1 в Китае. Число носителей гаплогруппы R1a1 у левитов — 51,7% (в 3 раза больше, чем у индийцев!). Это соответствует сербам (63,4%) и белорусам (51,0%). R1a1 (или по крайней мере R1a) имели протоиндоевропецы, протокартвелы, протоуральцы, протокавказцы, протодравиды и протоалтайцы.
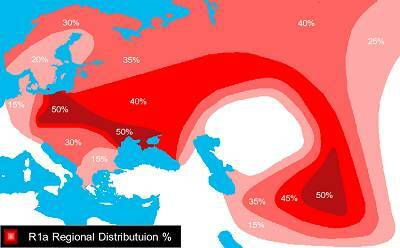
Карта распространения гаплогруппы R1a
Как объяснить столь широкую распространенность R1a1 за пределами индоевропейского ареала. Более того, остается загадкой, почему у левитов, киргизов, таджиков и других неиндоевропейских народов процент R1a1 больше, чем у самих индоевропейских народов, западных или средиземноморских?
Генетические исследования свидетельствуют о связи R1a1 с ностратической общностью
Как мы уже знаем, местом расселения «арийских» индусов являлся и является преимущественно север Индии. Однако, максимум вариации гаплогруппы R1a1 находится на западе Индостана, в низовьях Инда. Таким образом, следуя стандартному пути генетических расчетов, мы должны были бы сделать вывод, что эта область была локальной прародиной для индийских R1a1 (не для всех вообще, а именно для индийской подгруппы).
Конечно, для того, чтобы сделать вывод, где выше дисперсия, нужно на одинаковом уровне исследовать различные регионы. И чем больше регионов будет вовлечено в генетические исследования, включая Центральную Азию и Европу, тем яснее будет картина.
Напомним еще раз, что опубликованные в 2009 г. исследования С. Шармы показали индийское происхождение линии R1a1 среди брахманов. Еще в 2005 г. Сенгупта в своих исследованиях пришел к выводу, что R1a1 были у населения Индии гораздо раньше арийского вторжения. Сенгупта утверждает, что «миграции в период раннего голоцена (8000 — 10 000 лет назад) из северо-западной Индии (включая долину реки Инд) передали R1a1 (M17) как центральноазиатским, так и южноазиатским племенам». О древней миграции населения с Y-гаплогруппой R1a1 (M17) из Южной Азии в Европу говорят и исследования Питера Андерхилла.
Таким образом, напрашивается вывод о расселении носителей R1a1 задолго до начала экспансии «индоевропейцев» или «ариев», и все это наводит на мысль о связи R1a1 с ностратической общностью.
В эпоху палеолита численность населения была малой, и общность языка могла быть только генетической. Смена языка — явление, связанное с периодами формирования государственных или протогосударственных образований (союзов племен и т.п.), что для палеолита кажется невероятным. Существование языковой общности можно объяснить лишь расселением людей из компактного региона.
Все это может указывать на то, что маркеры R1a были у пра-ностратиков, и с их расселением они разошлись по всей Евразии. Возможно, произошел некий природный катаклизм, некое космическое явление или изменение магнитного поля Земли — и вот вам мутация, которая затронула большие группы людей.
Можем ли мы всерьёз предполагать, что некая индивидуальная мутация настолько изменила природу одного счастливого мальчика, что он затем разнёс свою изменённую хромосому по всему племени? Нет. Что-то значительное воздействовало на значимую группу мужских особей, из-за чего и происходило их выделение из прежней общины.
Продолжение см. в след. статье:
© П. Олексенко, 2011
См. еще:
С. А. Старостин. У человечества был единый праязык
Хараппская цивилизация оказалась древнейшей на Земле
Ведантист. Древнейшие города планеты. Самый древний город

1 comment
Leave a reply
Для отправки комментария вам необходимо авторизоваться.
Спасибо. Интересно.
Может такая карта полезной будет: https://glav.su/files/messages/2021/11/04/6209872_048dd22273480d2af79b1d3af3812e13.jpg
Оценка доли прото-индоевропейцев в современных жителях Европы